Research
○タンパク質膜輸送
細胞には生体膜で囲われた複雑な膜系が存在します。細胞を取り囲む細胞質膜だけでなく、小胞体やゴルジ体、ミトコンドリアなど、オルガネラ(細胞内小器官)もすべて生体膜で区画されています。これらのオルガネラで実際に作用している酵素類の多くはタンパク質であり、各オルガネラの機能発現に寄与しています。また、生体膜の中にも多くのタンパク質が局在しています。さらには、消化酵素やリゾチームなど、細胞の外にまで分泌して機能するタンパク質もたくさんあります。
ほとんどのタンパク質は細胞質で合成されます。しかし、多くのタンパク質が合成の場である細胞質以外に局在して機能を発揮しています。これらのタンパク質は、生体膜に組み込まれたり生体膜を横切って輸送されるわけです。生体膜は細胞の外と内を隔てる障壁となっているので、どんな物質でも自由に行き来できるようになっては困ったことになってしまいます。実際、ナトリウムイオンやカリウムイオン、糖類などの低分子化合物ですら生体膜の横断は厳しく制御されています。ましてや、タンパク質などの高分子は、自発的には膜挿入、膜透過はできなくなっているのです。そのため、タンパク質が生体膜を透過したり膜挿入したりするための「しくみ」が存在するはずです。さらには、膜輸送されるタンパク質とそうでないタンパク質も見分けなければなりませんし、目的地が異なるタンパク質もうまく仕分けなければなりません。こうしたタンパク質膜輸送に関わる課題はについては世界中で精力的に研究がすすめられてきました。
初期の研究では、分泌タンパク質がどのようにして膜を透過するのかについて研究がなされました。実は、分泌タンパク質のN末端には、「シグナルペプチド」と呼ばれる延長配列が付加されています。シグナルペプチドは、後に続くタンパク質を分泌させるための荷札のような働きをします。分泌タンパク質が合成されはじめると、SRP
(Signal recognition particle)がシグナルペプチドを認識し、SRP受容体(SR)を介して小胞体膜にまで導きます。その後シグナルペプチドが切断され、膜透過チャンネルを通ってタンパク質膜透過が進行するわけです。これが「シグナル仮説」と呼ばれるものです。(ただし、今では仮説ではありません。)分泌タンパク質が膜透過するしくみは全生物で同じ機構で起こると考えられました。バクテリア起源のベータラクタマーゼは小胞体を膜透過しますし、哺乳類のペプチドホルモン、プレプロラクチンは大腸菌でも膜透過するからです。さらに、SRPやSRはバクテリア、動植物を問わずすべての生物で発現していますし、タンパク質膜透過チャンネルは、SecYEG(バクテリア)、Sec61(真核生物)と名前こそ違いますが構造が似ている因子が発現しています。
膜タンパク質の膜挿入機構も同様に、合成途中の膜タンパク質がSRPにより認識され、SRを介して膜に導かれ、膜透過チャンネルにおいて膜挿入が進行していきます。図1は大腸菌の細胞質膜(内膜)と真核生物の小胞体膜へのタンパク質膜挿入の様子を簡単に図示したものですが、ほとんど同じしくみで膜挿入するといえます。YidCやTRAMは挿入途中の膜タンパク質とよく相互作用することが知られていますが、これらの機能を考えても膜挿入機構がよく保存されていることがわかります。
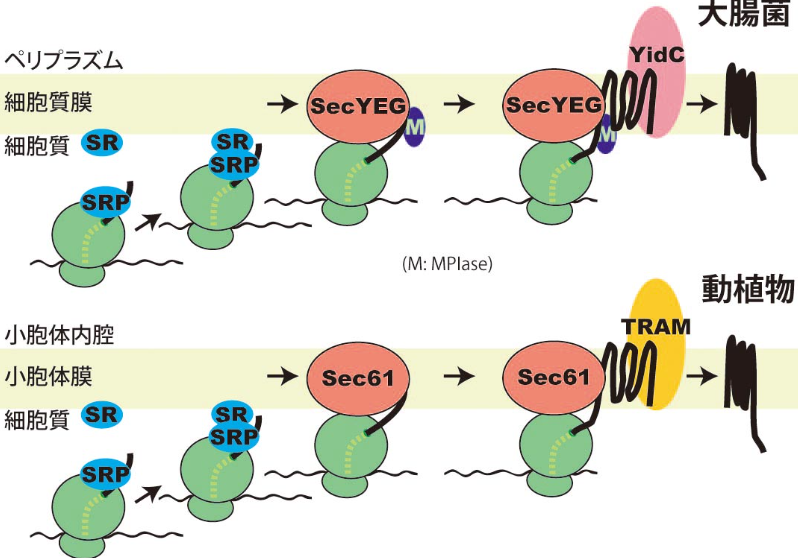
図1 大腸菌の細胞質膜(上)と真核生物の小胞体膜(下)への膜挿入機構
我々は、タンパク質膜透過反応や膜挿入反応の分子機構をより詳しく調べていくことを目的に研究を進めています。膜透過や膜挿入は細胞が生きていく上で不可欠な生命現象ですから、これらの反応に欠陥をもった変異体を調べるという手法ではどうしても限界があります。そのため、反応に必要な因子をすべて同定・精製して、それを試験管内で再構成して調べていく手法が有効です。こうした生化学的な研究を行うことにより、定量的で精度の高い研究ができると考えています。大腸菌はヒトに次いで研究されているモデル生物であり、多くの変異体が利用できるという点や培養が容易で実験材料も得やすいという点で優れた研究材料であると言えます。我々は大腸菌を用いて、動植物を問わずすべての生物に適用できるような普遍的な概念を導き出すことを目的にしています。
○大腸菌
「大腸菌」と聞くと、大腸の中の主要な細菌と思われるかもしれませんが、実は大腸の細菌の0.01~0.1%にしかならないマイナーな細菌です。腸内細菌の多くは酸素に触れると死滅してしまうので、好気呼吸ができる大腸菌が容易に分離・同定されたという歴史があります。また、病原性大腸菌が社会問題になりますが、もともとの大腸菌には病原性はありません。さらに、研究で用いられる大腸菌はいろんな変異やマーカーが入っているため、過酷な自然界ではまず生存競争に勝ち抜けないでしょう。
大腸菌は単細胞生物ですが、細胞表層構造は少し複雑です(図2)。「グラム陰性細菌」に分類され、外膜、内膜(細胞質膜)の2重の膜構造をもっています。外膜の内側にはペプチドグリカンがあり、細胞の骨格になっています。外膜と内膜の間には、「ペリプラズム空間」があり、高濃度の可溶性タンパク質が存在します。
大腸菌の染色体DNAは約465万塩基対あり、4千あまりのタンパク質をコードすると考えられています。その中で、細胞質に局在するタンパク質は約7割の3千弱しかありません。残りの3割はタンパク質生合成の場である細胞質ではなく、外膜、ペリプラズム、内膜に輸送されています。一部の病原性大腸菌を除けば外膜を超えてまで放出されるタンパク質はありません。外膜、ペリプラズムに局在するタンパク質の種類は比較的少ないですが、一種あたりの発現量が非常に多いタンパク質があるため、発現レベルを比べたとき、外膜、ペリプラズム、内膜にそれぞれ10%ずつになります。そのため、タンパク質の種類、発現量どちらをとっても約30%のタンパク質は細胞質以外の場所に輸送されているわけです。
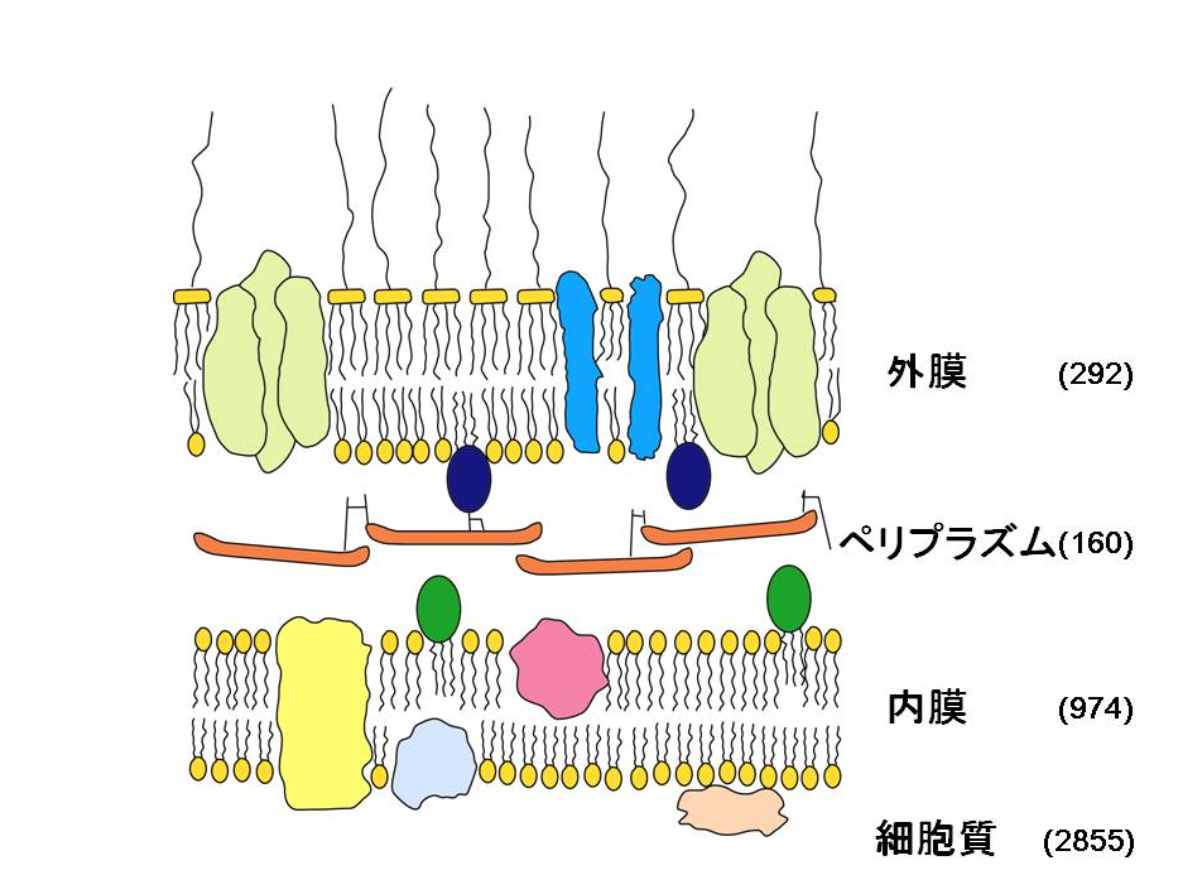
図2 大腸菌の細胞表層構造。数字はそれぞれの場所に局在すると考えられているタンパク質の種類
○タンパク質膜透過
さて、大腸菌の外膜とペリプラズムに局在するタンパク質は、細胞質で合成された後、内膜を透過しなければなりません。そのため、これらのタンパク質は、上述の分泌タンパク質同様、N末端にシグナルペプチド(リーダーペプチド)をもった前駆体タンパク質として合成されます。大腸菌では通常外膜を超えてタンパク質が放出されることはありませんが、その構造的類似性から、ペリプラズムタンパク質も外膜タンパク質も「分泌タンパク質」と定義されています。動物由来の分泌タンパク質は大腸菌の内膜を透過しますし、バクテリア由来の分泌タンパク質が小胞体膜を透過したりして、タンパク質膜透過機構はどんな生物でもよく似たしくみで進行すると考えられてきました。実際、SecYEGとSec61はよく似ていてどちらも分泌タンパク質が透過していく「ポア」になりますし、SRP、SRはどんな生物にも存在します。
しかし、全く同じ機構でタンパク質膜透過が起こるかどうかというと、実はかなり違います。その大きな違いは、小胞体膜透過はタンパク質合成に共役して進行する(co-translational)のに対して、大腸菌ではタンパク質合成が大部分終了してから(post-translational)膜透過する点です。そのため、大腸菌ではSRPはタンパク質膜透過には関わらないわけです。SRPは「シグナル認識粒子」だったはずで、大腸菌ではSRPはシグナルペプチドを認識しないというのはとても紛らわしいですが、SRPには実はもっと重要な仕事があります。それは、タンパク質膜透過ではなく、タンパク質膜挿入です。
○タンパク質膜挿入
膜タンパク質は膜に局在するタンパク質ですが、その中でも「膜内在性タンパク質」は膜を貫通する領域をもっています。この領域はとても疎水的(あぶらっこい)ので、水には溶けません。この疎水的な領域が合成されると、速やかに手当てしないと凝集体ができてしまいます。そのため、膜タンパク質は合成されると同時に膜に挿入されます。膜タンパク質が合成され始め、疎水的な膜貫通領域がリボソームから出てくるとそこにSRPが結合します。SRPが結合した合成途中の膜タンパク質は膜上のSRP受容体(SR)に輸送されます。その後、膜透過にも必要なSecYEG上で膜挿入が進行します。YidCは膜挿入途中のタンパク質と相互作用し、おそらく膜タンパク質が膜内で高次構造を形成するのに必要です。図3の上段は、動植物の小胞体膜への膜挿入に驚くほどよく似ていることがおわかりいただけると思います。
これだけで全てが説明できれば非常にすっきりとするのですが、実際にはもう少しバリエーションがあります。もし、膜タンパク質が図3中段のように親水的な領域をペリプラズム側にもっていれば、SecAという因子がさらに必要です。SecAはタンパク質膜透過に必要な因子でATPase活性を持っています。親水的な領域は、分泌タンパク質の膜透過と同様にSecAにより膜透過されます。しかし、この場合も途中までは上段と同じです。
さらにややこしいのは、下段の「Sec非依存」の経路の存在です。この経路ではSecだけでなくSRPも不要です。なぜかというと、この経路の膜タンパク質は分子量が小さいものが多く、SRPが膜貫通領域に相互作用しようとしたときにはすでにタンパク質合成が終了してしまっているからです。SRPは合成途中のタンパク質にしか結合できないのです。さらに、この経路の膜タンパク質は、リン脂質だけでできたリポソーム(人工的な膜小胞)に膜挿入することが知られています。したがって、Sec非依存の膜タンパク質は、膜脂質と膜タンパク質の疎水性相互作用のみによって「自発的に膜挿入」すると考えられてきました。
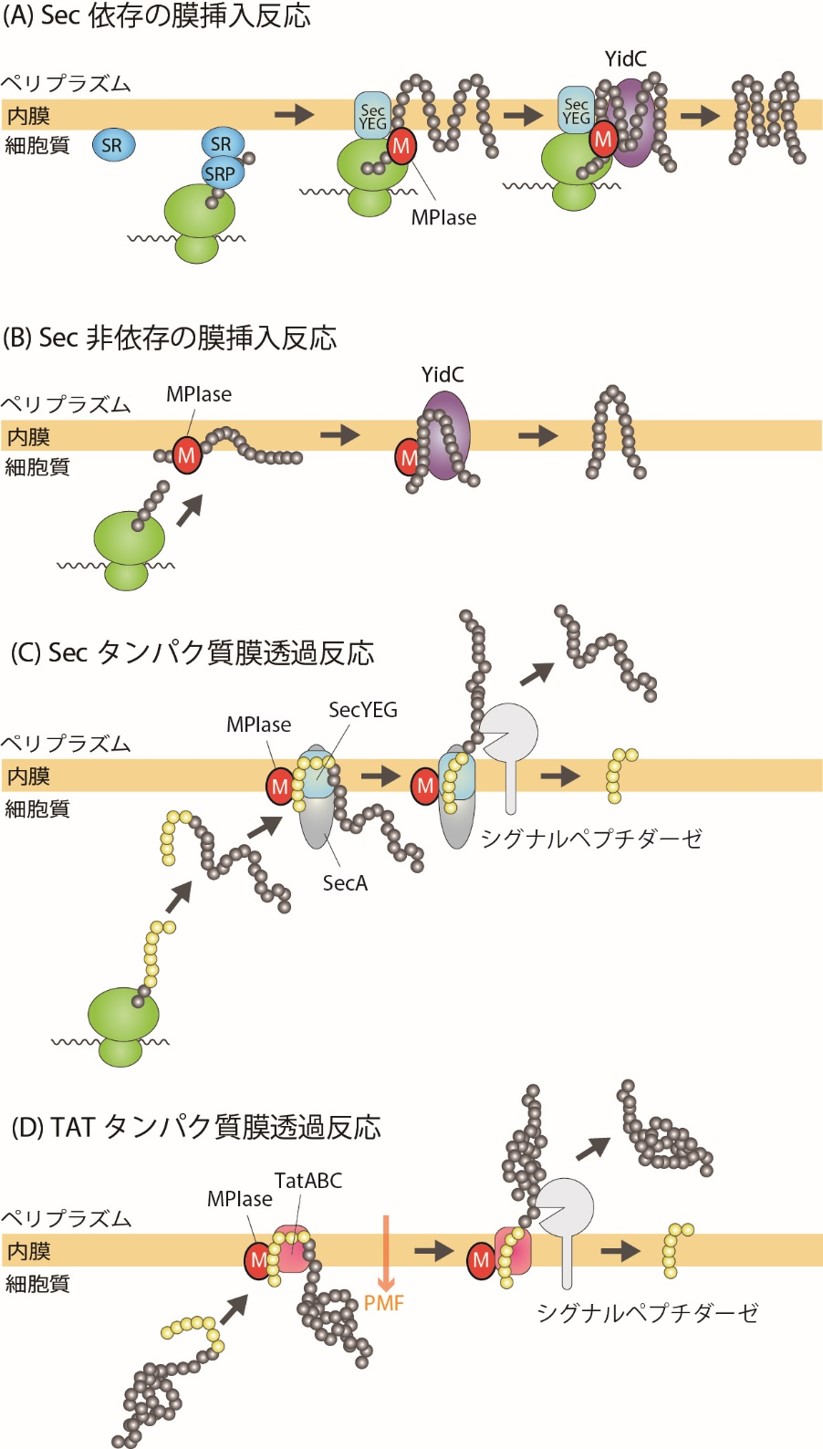
図3 大腸菌におけるタンパク質膜挿入機構
○自発的膜挿入とジアシルグリセロール
「Sec非依存」の膜タンパク質は本当に自発的膜挿入するのでしょうか?これについてはかなり強力な証拠があります。まず、プロテアーゼを封入したリポソームを作製します。このリポソーム存在下でSec非依存の膜タンパク質を合成させると、膜挿入が起こり、部分的に膜透過した部分がリポソーム内部のプロテアーゼによって分解されてしまうのです。
ところで、私がタンパク質膜挿入の研究を始めたのは、MtlA(マンニトール・パーミアーゼ)という膜タンパク質の膜挿入を再構成を目指したときです。MtlAは図3の上段のような機構で膜挿入することが知られていました。そのため、SecYEGやYidC、SRPとSRを用意し、リポソームにこれらを組み込んでMtlAが膜挿入するかどうか調べました。このとき、コントロールを取るためこれらの因子を全く組み込んでいないリポソームも用意しました。実験してみて驚いたことに、MtlAはこのリポソームにも膜挿入したのです。一方、SecYEGを枯渇させた膜小胞では、MtlAは全く膜挿入しません。このことから、リポソームへのMtlAの膜挿入は、生理的な状態を全く反映していないと結論せざるを得なくなりました。
では、なぜMtlAは因子を何も組み込んでいないリポソームに膜挿入するのでしょうか?私は、リポソームがきれいすぎるのではないかと考えました。生化学の研究では、因子を徹底的に精製してから研究を行います。不純物によるバイアスを防ぐためにもとても重要なことです。しかし、リポソームは生体膜と比較すれば単純すぎるのかもしれません。さらに考えを進めると、そもそも膜は疎水的なので自発的膜挿入というのは不可避な現象であって、生体膜は無秩序な自発的膜挿入を防ぐために積極的に策を講じているのではないかということになります。そうすると、生体膜には自発的膜挿入を防ぐための物質が存在するはずです。このような仮説のもと、生体膜を構成する成分をいろいろと調べました。その結果、ジアシルグリセロールが自発的膜挿入をブロックすることを見い出しました。ジアシルグリセロールは大腸菌の膜には1~2%存在する因子であり、リポソームにも同じくらい加えてやるとMtlAの自発的膜挿入は全く起こらなくなりました。
Sec非依存の膜タンパク質の膜挿入もジアシルグリセロール存在下で調べてみると、MtlAと同様に全く膜挿入しなくなることがわかりました。したがって、リン脂質にはどんな膜タンパク質も自発的膜挿入することができるが、これは生理的な膜挿入を反映しているわけではなく、実際細胞の中ではジアシルグリセロールによって自発的膜挿入は起こらない仕組みになっているということが言えます。
○糖脂質酵素MPIase
MtlAの自発的膜挿入はジアシルグリセロールにより完全にブロックすることができました。では、ジアシルグリセロールを含むリポソームにSecYEGやYidC、SRPとSRを組み込めばMtlAは膜挿入するでしょうか?答えは「膜挿入しない」でした。この結果は、膜挿入に必要な因子はこれだけではないということを意味します。そういえば、Sec非依存の膜タンパク質も、ジアシルグリセロールが存在すれば自発的膜挿入はしないと述べました。では、何に依存して膜挿入するのでしょうか。SecYEGやYidC、SRPがあっても膜挿入はしませんでした。このことも何か別の因子が存在しているということを示しています。
タンパク質膜挿入に関わる新因子があるということになると、これを精製・同定しないといけません。私はこの手の実験が好きですが、そもそも微量の因子なので大変でした。まず、本当に新因子があるのかどうか示すため、大腸菌の膜画分をいくつかの画分に分けました。この中の一つが、既知の膜挿入因子(SRPやYidC、Sec因子など)を全く含まないけれども、SecYEGと混ぜてリポソームに組み込むと活性があることがわかりました。この画分を予めプロテアーゼ処理しておくと膜挿入活性がなくなるので、何かタンパク質性の因子が働いていると考えました。また、この画分を組み込んだリポソームでは、Sec非依存の膜挿入活性も検出できることがわかりました。したがって、タンパク質膜挿入一般に働く新因子が存在するということです。さらに精製を進めていきました。結果的に、電気泳動をすると約8
kDaの位置に膜挿入活性のピークと一致するバンドが得られました。これを「MPIase(Membrane Protein Integrase)」と呼ぶことにしました。
さて、MPIaseがタンパク質であれば、その配列は遅かれ早かれ確定するはずです。しかし、そのアミノ酸配列の情報は全く得られませんでした。それもそのはず、MPIaseはタンパク質ではなかったのです。最近、MPIaseの全構造が明らかになりました(図4)。それによると、MPIaseは複合糖脂質でした。ただし、糖鎖部分にはアセチル基がたくさん(1分子あたり約35個)ありました。アセチル基の大部分はN-アセチル基でした。アセチル基はMPIase機能に必須なので、プロテアーゼでN-アセチル基が切断されて失活したのかもしれません。また、MPIaseは8量体を形成している可能性も明らかになってきました。そのため、リボソームで合成された直後の膜タンパク質がMPIaseに取り囲まれ、その後膜挿入反応が進行すると考えられます(図5)。膜タンパク質の疎水的な領域とMPIaseのアセチル基が疎水的相互作用していると考えられます。したがって、MPIaseは酵素のような働きをもっている糖脂質であるといえます。このことから、MPIaseが「糖脂質酵素(Glycolipozyme)」と呼ぶべきではないかと提唱しています(Nat
Commun, 2012)。
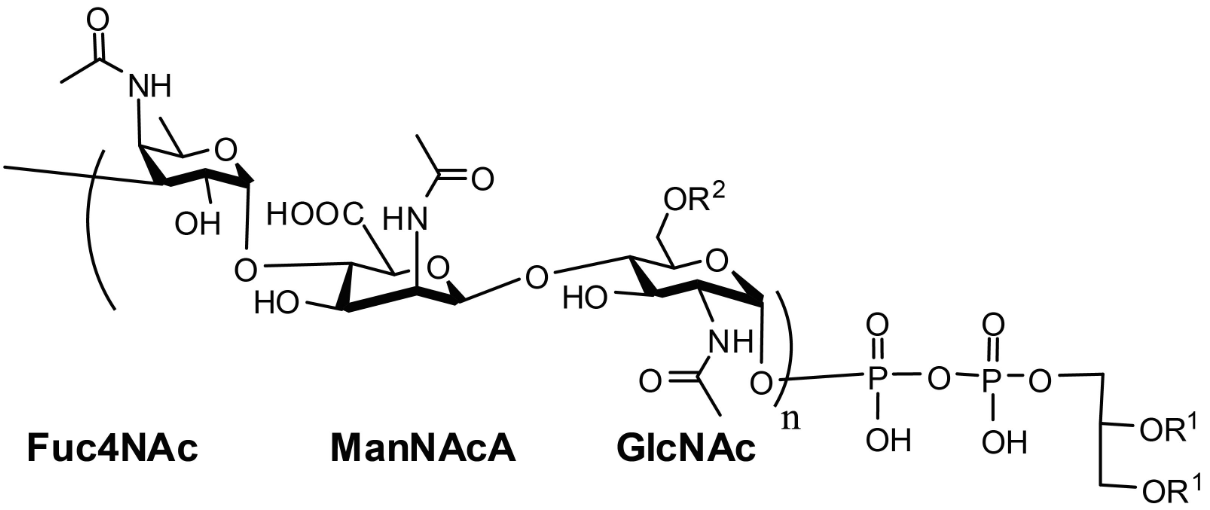
図4 MPIaseの構造。GlcNAcはNアセチルグルコサミン、ManNAcAはNアセチルマンヌロン酸、Fuc4NAcは4デオキシ4アセトアミドフコースを示す。R1はC16~C20の脂肪酸、R2はアセチル基またはOH基を示す。nは9~11の整数を示す。
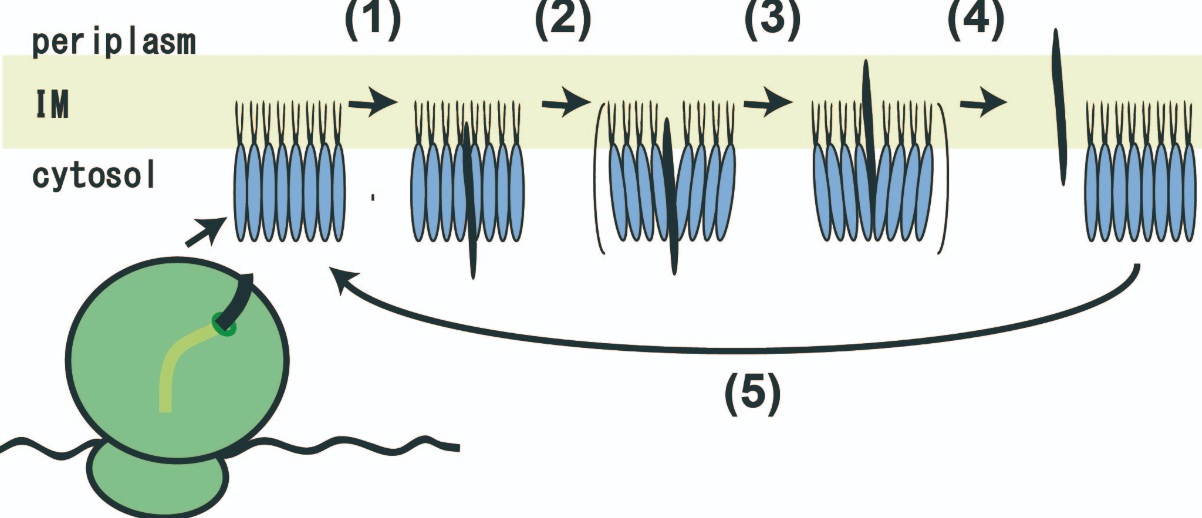
図5 MPIaseに依存したタンパク質膜挿入。
○MPIaseとSecYEGとの相互作用
話は変わりますが、我々は膜透過装置SecYEGのうち、SecGを同定しました(EMBO J 1993, 1994)。さらにSecGの研究を進めていくと、SecGは図6のような構造変化をすることがわかりました(Cell
1996; EMBO J 1999; J Biol Chem 2007; 2012など)。SecAはATPase活性をもっているタンパク質で、ATPを加水分解しながら分泌タンパク質を細胞質からペリプラズムに押し出す働きをもっています。このピストンのようなSecAの構造変化に共役しながらSecGが構造変化するわけですが、そのとき膜を2回貫通するSecGは反転-回復を繰り返すわけです。膜タンパク質というのは予めその膜内配向性がアミノ酸配列によって決定されており、それは生涯変化することはないとされてきました。そのため、SecGの配向性反転は、他に例を見ない画期的な概念だったわけです。さて、最近明らかになったのは、SecG反転サイクルにはMPIaseが必須であることでした(PNAS,
2013)。別々の研究が一つに繋がった珍しいケースといえます。
SecYEGを組み込んだリポソームにMPIaseを共存させると、SecYEGしかないときに比べて膜透過反応が10倍以上促進されます。その理由は、SecYEGの二量体構造がMPIaseの有無で全く異なることでした。SecYEGはきれいなX線結晶構造が明らかになっていて、図7の左のような構造をしています。SecEが二量体の接触面にあり、「Back-to-back構造」と呼ばれています。それに対して、MPIaseが存在するとSecYEGの二量体構造が大きく変化して、図7右のような構造に変化します。我々はこれを「Side-by-side構造」と呼んでいます。SecYEGがSide-by-side構造を取るとSecG反転サイクルが作動できる状態になり、その結果、膜透過反応が著しく上昇すると考えられます。結晶を作成するときは非常に純粋なSecYEGが必要となります。純粋な精製剽品を調製する際は、まずSecYEGを大腸菌で過剰生産します。このとき、MPIaseの量は変化しませんから、MPIaseの量が不足してしまいます。その後精製されたSecYEGにはMPIaseがほとんど含まれていないためにBack-to-back構造になってしまったのでしょう。MPIaseがさまざまな機能をもっていることがおわかりいただけると思います。
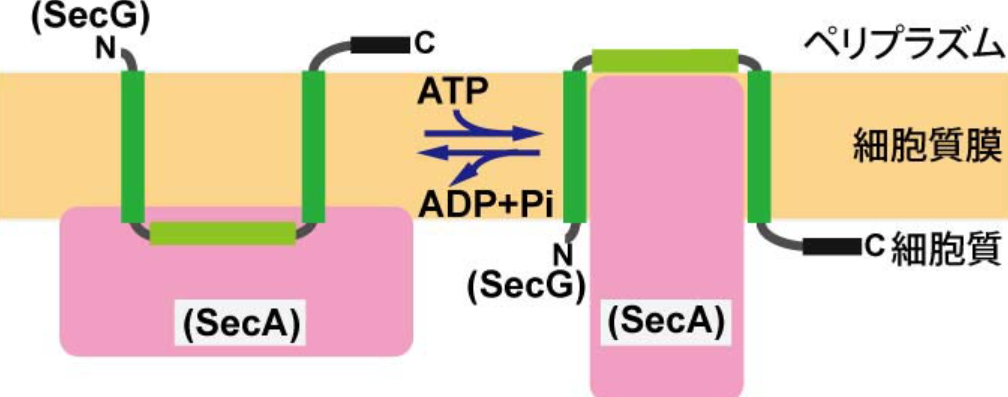
図6 タンパク質膜透過反応に共役したSecGの反転
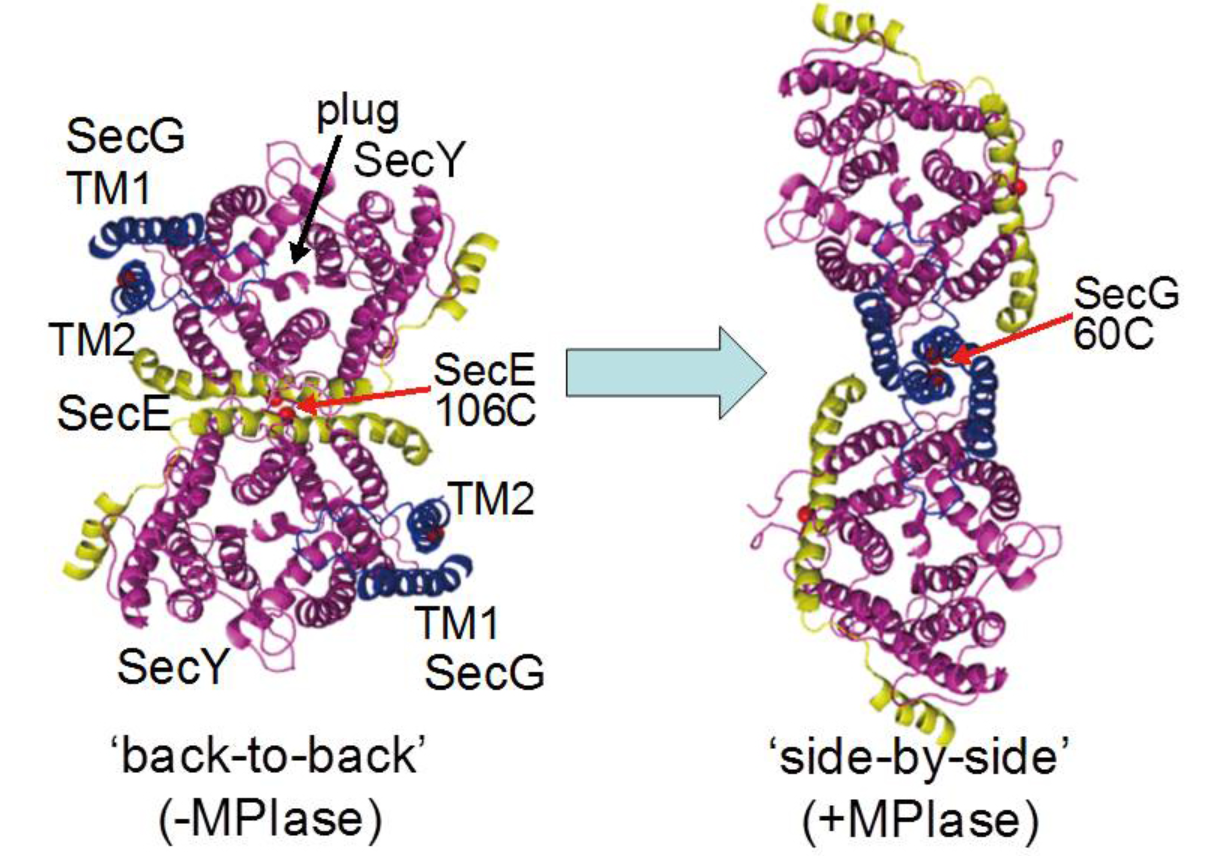
図7 MPIaseによるSecYEG二量体の構造変化