2025年4月の農学部組織改編に伴い、当研究室は「生命科学科・分子生命医科学コース・分子生物学研究室」となり、新たな気持ちで研究活動を開始することとなりました。これまで「寒冷バイオフロンティア研究センター・寒冷発育制御研究分野」や「応用生物化学科・分子生物学研究室」において展開してきた基礎研究をさらに発展させるとともに、新学科およびコースの目標である「QOL
(Quality of Life)の向上」に資する研究にも取り組んでまいります。
ご挨拶
研究概要
DNA の遺伝情報が mRNA に転写され、タンパク質に翻訳されるというのは分子生物学の「セントラル・ドグマ」と呼ばれるもので、遺伝子の発現の流れを示すものです。しかし、このとき生成するのは、厳密にはタンパク質ではなく、アミノ酸がペプチド結合で重合した鎖状の化合物、「ポリペプチド」に過ぎません。実際に「タンパク質」として機能するためには、ポリペプチドに適切な修飾が施され、高次構造が形成させるなどの過程を経なければなりませんし、さらにはそのタンパク質が実際に機能する場所に輸送されなければなりません。つまり、タンパク質は適材適所に配置される必要があるのです。我々はそのしくみを研究しています。
そのしくみが解明されれば、新しい薬剤の開発、老化のメカニズムの解明、低温に強い作物の開発などが期待できます。
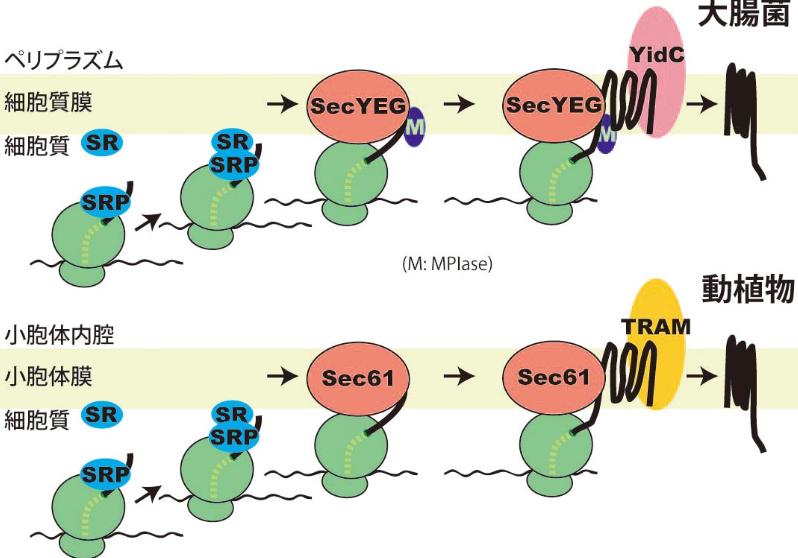
タンパク質は通常細胞質で生合成されますが、細胞小器官(オルガネラ)や細胞の外など、細胞質以外で機能するものもたくさんあります。こうしたタンパク質は生体膜を透過したり生体膜に組み込まれたりする必要があります。生体膜(特に細胞質膜)は外界と自己の境目になっていて、膜を介した物質の横断は厳密に制御されています。たとえば、イオンや糖類などの低分子はおろか水分子さえ膜を自由に行き来することはできません。したがって、タンパク質がこうした生体膜を透過したり組み込まれたりするためには巧妙な仕組みが必要となってきます。下図は膜タンパク質が動植物細胞の小胞体膜や大腸菌の細胞質膜にどうやって組み込まれるかを大まかに示したものです。一連の反応に必要な因子の名前こそ若干違いますが、その基本的な反応の流れはどちらも驚くほど類似しています。こうした生物全般に保存された生命現象を、モデル生物大腸菌を用いて詳細で精密な研究により明らかにしていこうというのが我々の研究目的です。
我々は、タンパク質膜透過反応や膜挿入反応に必要な因子をすべて純化し、試験管の中で再構成することに成功しました。さらには、タンパク質膜挿入反応に必要な糖脂質MPIaseも同定しました。この糖脂質はタンパク質膜挿入反応を触媒するという酵素のような機能をもつことから、MPIase(Membrane
Protein Integrase)と名付けました。現在はMPIaseの構造と機能の関係について調べています。
大腸菌における膜挿入反応と動植物における膜挿入反応がよく似ているということは、動植物においてもMPIaseに相当する物質が存在すると考えられます。また、植物の葉緑体では、大腸菌と同じようなタンパク質輸送が起きていることが知られています。最近、動物細胞や植物細胞でもMPIaseのような活性をもつ物質があることがわかってきました。現在、動物や植物のMPIaseの構造解析・機能解析も進めています。





_
     |
{kind=link}